Репродуктивная функция женщин осуществляется прежде всего благодаря деятельности яичников и матки, так как в яичниках созревает яйцеклетка, а в матке под влиянием гормонов, выделяемых яичниками, происходят изменения по подготовке к восприятию оплодотворенного плодного яйца. Репродуктивный период характеризуется способностью организма женщины к воспроизводству потомства; продолжительность данного периода от 17—18 до 45—50 лет. Репродуктивному, или детородному, периоду предшествуют следующие этапы жизни женщины: внутриутробный; новорожденный (до 1 года); детства (до 8—10 лет); препубертатного и пубертатного возраста (до 17—18 лет). Репродуктивный период переходит в климактерический, в котором различают пременопаузу, менопаузу и постменопаузу.
Менструальный цикл — одно из проявлений сложных биологических процессов в организме женщины. Менструальный цикл характеризуется циклическими изменениями во всех звеньях репродуктивной системы, внешним проявлением которых является менструация.
Менструации — это кровянистые выделения из половых путей женщины, периодически возникающие в результате отторжения функционального слоя эндометрия в конце двухфазного менструального цикла. Первая менструация (menarhe) наблюдается в возрасте 10—12 лет, но в течение 1 — 1,5 года после этого менструации могут быть нерегулярными, а затем устанавливается регулярный менструальный цикл.
Первый день менструации условно принимается за первый день менструального цикла. Следовательно, продолжительность цикла составляет время между первыми днями двух последующих менструаций. Для 60 % женщин средняя продолжительность менструального цикла составляет 28 дней с колебаниями от 21 до 35 дней. Величина кровопотери в менструальные дни 40—60 мл, в среднем 50 мл. Продолжительность нормальной менструации от 2 до 7 дней.
Яичники. Во время менструального цикла в яичниках происходит рост фолликулов и созревание яйцеклетки, которая в результате становится готовой к оплодотворению. Одновременно в яичниках вырабатываются половые гормоны, обеспечивающие изменения в слизистой оболочке матки, способной воспринять оплодотворенное яйцо.
Половые гормоны (эстрогены, прогестерон, андрогены) являются стероидами, в их образовании принимают участие фанулезные клетки фолликула, клетки внутреннего и наружного слоев. Половые гормоны, синтезируемые яичниками, влияют на ткани- и органы-мишени. К ним относятся половые органы, в первую очередь матка, молочные железы, губчатое вещество костей, мозг, эндотелий и гладкие мышечные клетки сосудов, миокард, кожа и ее придатки (волосяные фолликулы и сальные железы) и др. Прямой контакт и специфическое связывание гормонов на клетке-мишени является результатом взаимодействия его с соответствующими рецепторами.
Биологический эффект дают свободные (несвязанные) фракции эстрадиола и тестостерона (1 %). Основная масса яичниковых гормонов (99 %) находится в связанном состоянии. Транспорт осуществляется специальными белками — стероидосвязывающими глобулинами и неспецифическими транспортными системами — альбуминами и эритроцитами.
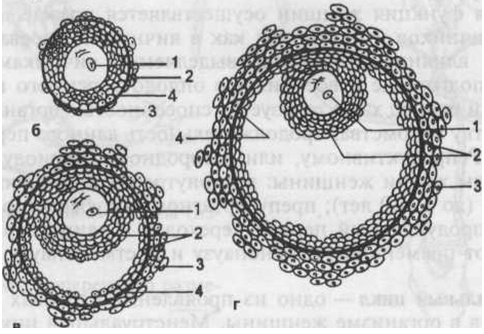
Рисунок: Этапы развития доминантного фолликула.
а — примордиальный фолликул; б — преантралъный фолликул; в — антральный фолликул; г — преовуляторный фолликул: 1 — овоцит, 2 — гранулезные клетки (зернистая зона), 3 — тека-клетки, 4 — базальная мембрана.
Эстрогенные гормоны способствуют формированию половых органов, развитию вторичных половых признаков в период полового созревания. Андрогены оказывают влияние на появление оволосения на лобке и в подмышечных впадинах. Прогестерон контролирует секреторную фазу менструального цикла, подготавливает эндометрий к имплантации. Половые гормоны играют важную роль в процессе развития беременности и родов.
Циклические изменения в яичниках включают три основных процесса:
1. Рост фолликулов и формирование доминантного фолликула.
Овуляция.
Образование, развитие и регресс желтого тела.
При рождении девочки в яичнике находятся 2 млн фолликулов, 99 % которых подвергаются атрезии в течение всей жизни. Под процессом атрезии понимается обратное развитие фолликулов на одной из стадий его развития. Ко времени менархе в яичнике содержится около 200—400 тыс. фолликулов, из которых созревают до стадии овуляции 300—400.
Принято выделять следующие основные этапы развития фолликула: примордиальный фолликул, преантральный фолликул, антральный фолликул, преовуляторный фолликул.
Примордиальный фолликул состоит из незрелой яйцеклетки, которая расположена в фолликулярном и гранулезном (зернистом) эпителии. Снаружи фолликул окружен соединительной оболочкой (тека-клетки). В течение каждого менструального цикла от 3 до 30 примордиальных фолликулов начинают расти, и из них формируются преантральные, или первичные, фолликулы.
Преантральный фолликул. С началом роста примордиальный фолликул прогрессирует до преантральной стадии, а овоцит увеличивается и окружается мембраной, называемой блестящей оболочкой (zona pellucida). Клетки гранулезного эпителия подвергаются размножению, а слой теки образуется из окружающей стромы. Этот рост характеризуется повышением продукции эстрогенов. Клетки гранулезного слоя преантрального фолликула способны синтезировать стероиды трех классов, при этом эстрогенов синтезируется гораздо больше, чем андрогенов и прогестерона.
Антральный, или вторичный, фолликул . Характеризуется дальнейшим ростом: увеличивается число клеток гранулезного слоя, продуцирующих фолликулярную жидкость. Фолликулярная жидкость накапливается в межклеточном пространстве гранулезного слоя и образует полости. В этот период фолликулогенеза (8—9-й день менструального цикла) отмечается синтез половых стероидных гормонов, эстрогенов и андрогенов.
Согласно современной теории синтеза половых гормонов, в тека-клетках синтезируются андрогены — андростендион и тестостерон. Затем андрогены попадают в клетки гранулезного слоя, и в них ароматизируются в эстрогены.
Доминантный фолликул. Как правило, один такой фолликул образуется из множества антральных фолликулов (к 8-му дню цикла). Он является самым крупным, содержит наибольшее число клеток гранулезного слоя и рецепторов к ФСГ, ЛГ. Доминантный фолликул имеет богато васкуляризированный тека-слой. Наряду с ростом и развитием доминантного преовуляторного фолликула в яичниках параллельно происходит процесс атрезии остальных (90 %) растущих фолликулов.
Доминантный фолликул в первые дни менструального цикла имеет диаметр 2 мм, который в течение 14 дней к моменту овуляции увеличивается в среднем до 21 мм. За это время происходит 100-кратное увеличение объема фолликулярной жидкости. В ней резко возрастает содержание эстрадиола и ФСГ, а также определяются факторы роста.
Овуляция— разрыв преовулярного доминантного (третичного) фолликула и выход из него яйцеклетки. Ко времени овуляции в овоците происходит процесс мейоза. Овуляция сопровождается кровотечением из разрушенных капилляров, окружающих тека-клетки. Полагают, что овуляция происходит через 24—36 ч после формирования преовуляторного пика эстрадиола. Истончение и разрыв стенки преовуляторного фолликула происходят под влиянием фермента коллагеназы. Определенную роль играют также простагландины F2a и Ег, содержащиеся в фолликулярной жидкости; протеолитические ферменты, образующиеся в гранулезных клетках; окситоцин и релаксин.
После выхода яйцеклетки в полость фолликула быстро врастают образующиеся капилляры. Гранулезные клетки подвергаются лютеинизации: в них увеличивается объем цитоплазмы и образуются липидные включения. ЛГ, взаимодействуя с белковыми рецепторами гранулезных клеток, стимулирует процесс их лютеинизации. Этот процесс приводит к образованию желтого тела.
Желтое тело — транзиторная эндокринная железа, которая функционирует в течение 14 дней независимо от продолжительности менструального цикла. При отсутствии беременности желтое тело регрессирует.
Таким образом, в яичнике синтезируются основные женские половые стероидные гормоны — эстрадиол и прогестерон, а также андрогены.
В I фазу менструального цикла, который длится от первого дня менструации до момента овуляции, организм находится под влиянием эстрогенов, а во II (от овуляции до начала менструации) к эстрогенам присоединяется прогестерон, выделяющийся клетками желтого тела. Первая фаза менструального цикла называется также фолликулиновой, или фолликулярной, вторая фаза цикла — лютеиновой.
В течение менструального цикла в периферической крови отмечаются два пика содержания эстрадиола: первый — выраженный преовуляторный цикл, и второй, менее выраженный, — в середине второй фазы менструального цикла. После овуляции во второй фазе цикла основным является прогестерон, максимальное количество которого синтезируется на 4—7-й день после овуляции.
Циклическая секреция гормонов в яичнике определяет изменения в слизистой оболочке матки.
Циклические изменения в слизистой оболочке матки (эндометрии).Эндометрий состоит из следующих слоев:
Базальный слой, который не отторгается во время менструации. Из его клеток в течение менструального цикла образуется слой эндометрия.
Поверхностный слой, состоящий из компактных эпителиальных клеток, которые выстилают полость матки.
Промежуточный, или спонгиозный, слой.
Последние два слоя составляют функциональный слой, подвергающийся основным циклическим изменениям в течение менструального цикла и отторгающийся в период менструации.
В I фазе менструального цикла эндометрий представляет собой тонкий слой, состоящий из желез и стромы. Выделяют следующие основные фазы изменения эндометрия в течение цикла:
1) фаза пролиферации;
2) фаза секреции;
3) менструация.
Фаза пролиферации. По мере увеличения секреции эстрадиола растущими фолликулами яичников эндометрий претерпевает пролиферативные изменения. Происходит активное размножение клеток базального слоя. Образуется новый поверхностный рыхлый слой с вытянутыми трубчатыми железами. Этот слой быстро утолщается в 4—5 раз. Трубчатые железы, выстланные цилиндрическим эпителием, удлиняются.
Фаза секреции. В лютеиновую фазу яичникового цикла под влиянием прогестерона увеличивается извилистость желез, а просвет их постепенно расширяется. Клетки стромы, увеличиваясь в объеме, приближаются друг к другу. Секреция желез усиливается. В просвете желез находят обильное количество секрета. В зависимости от интенсивности секреции железы либо остаются сильно извитыми, либо приобретают пилообразную форму. Отмечается усиленная васкуляризация стромы. Различают раннюю, среднюю и позднюю фазы секреции.
Менструация. Это отторжение функционального слоя эндометрия. Тонкие механизмы, лежащие в основе возникновения и процесса менструации, неизвестны. Установлено, что эндокринной основой начала менструации является выраженное снижение уровней прогестерона и эстрадиола вследствие регрессии желтого тела.
Существуют следующие основные локальные механизмы, принимающие участие в менструации:
1) изменение тонуса спиральных артериол;
2) изменение механизмов гемостаза в матке;
3) изменения в лизосомной функции клеток эндометрия;
4) регенерация эндометрия.
Установлено, что началу менструации предшествует интенсивное сужение спиральных артериол, приводящее к ишемии и десквамации эндометрия.
В течение менструального цикла изменяется содержание лизосом в клетках эндометрия. Лизосомы содержат ферменты, некоторые из которых участвуют в синтезе простагландинов. В ответ на снижение уровня прогестерона усиливается выделение этих ферментов.
Регенерация эндометрия наблюдается с самого начала менструации. К концу 24-го часа менструации отторгается 2/з функционального слоя эндометрия. Базальный слой содержит эпителиальные клетки стромы, являющиеся основой для регенерации эндометрия, которая обычно к 5-му дню цикла полностью завершается. Параллельно завершается ангиогенез с восстановлением целости разорванных артериол, вен и капилляров.
Изменения в яичниках и матке происходят под влиянием двухфазной деятельности регулирующих менструальную функцию систем: кора большого мозга, гипоталамус, гипофиз. Таким образом, выделяются 5 основных звеньев репродуктивной системы женщины: кора большого мозга, гипоталамус, гипофиз, яичник, матка. Взаимосвязь всех звеньев репродуктивной системы обеспечивается наличием в них рецепторов как к половым, так и гонадотропным гормонам.
О роли ЦНС в регуляции функции репродуктивной системы известно давно. Об этом свидетельствовали нарушения овуляции при различных острых и хронических стрессах, нарушение менструального цикла при перемене климатогеографических зон, ритма работы; хорошо известно прекращение менструаций в условиях военного времени. У психически неуравновешенных женщин, страстно желающих иметь ребенка, менструации также могут прекратиться.
В коре большого мозга и в экстрагипоталамических церебральных структурах (лимбической системе, гиппо-кампе, миндалине и др.) выявлены специфические рецепторы для эстрогенов, прогестерона и андрогенов. В этих структурах происходят синтез, выделение и метаболизм нейропептидов, нейротрансмиттеров и их рецепторов, которые в свою очередь избирательно влияют на синтез и выделение рилизинг-гормона гипоталамуса.
Во взаимосвязи с половыми стероидами функционируют нейротрансмиттеры: норадреналин, дофамин, гамма-аминомасляная кислота, ацетилхолин, серотонин и мелатонин. Норадреналин стимулирует выброс гонадотропин-рилизинг-гормона (ГТРГ) из нейронов переднего гипоталамуса. Дофамин и серотонин уменьшают частоту и снижают амплитуду выработки ГТРГ в различные фазы менструального цикла.
Нейропептиды (эндогенные опиоидные пептиды, нейропептид Y, кортикотропин-рилизинг-фактор и галанин) также влияют на функцию репродуктивной системы, а следовательно, на функцию гипоталамуса. Эндогенные опиоидные пептиды трех видов (эндорфины, энкефалины и динорфины) способны связываться с опиатными рецепторами мозга. Эндогенные опиоидные пептиды (ЭОП) модулируют влияние половых гормонов на содержание ГТРГ по механизму обратной связи, блокируют секрецию гипофизом гонадотропных гормонов, особенно ЛГ, посредством блокады секреции ГТРГ в гипоталамусе.
Взаимодействие нейротрансмиттеров и нейропептидов обеспечивает в организме женщины репродуктивного возраста регулярные овуляторные циклы, влияя на синтез и выделение ГТРГ гипоталамусом.
В гипоталамусе имеются пептидергические нейронные клетки, которые секретируют стимулирующие (либерины) и блокирующие (статины) нейрогормоны — нейросекреция. Эти клетки обладают свойствами как нейронов, так и эндокринных клеток, и отвечают как на сигналы (гормоны), поступающие из кровотока, так и на нейротрансмиттеры и нейропептиды мозга. Нейрогормоны синтезируются в рибосомах цитоплазмы нейрона, а затем транспортируются по аксонам к терминалям.
Гонадотропин-рилизинг-гормон (либерин) — нейрогормон, регулирующий гонадотропную функцию гипофиза, где синтезируются ФСГ и ЛГ. Рилизинг-гормон ЛГ (люлиберин) выделен, синтезирован и подробно описан. Выделить и синтезировать рилизинг-фолликулостимулирующий гормон, или фоллиберин, до настоящего времени не удалось.
Секреция гонадолиберина имеет пульсирующий характер: пики усиленной секреции гормона продолжительностью несколько минут сменяются 1— 3-часовыми интервалами относительно низкой секреторной активности. Частоту и амплитуду секреции гонадолиберина регулирует уровень эстрогенов.
Нейрогормон, который контролирует секрецию пролактина аденогипофизом, называется пролактинингибирующим гормоном (фактором), или дофамином.
Важным звеном в системе репродукции является передняя доля гипофиза — аденогипофиз, в котором секретируются гонадотропные гормоны, фолликулостимулирующий гормон (ФСГ, фоллитропин) лютеинизирующий гормон (ЛГ, лютропин) и пролактин (Прл), регулирующие функцию яичников и молочных желез. Все три гормона являются белковыми веществами (полипептидами). Железой-мишенью гонадотропных гормонов является яичник.
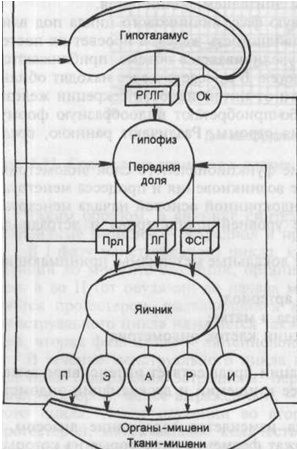
Рисунок: Функция репродуктивной системы (схема).
РГЛГ — рилизинг-гормоны; ОК — окситоцин; Прл — пролактин; ФСГ — фолликуло-стимулирующий гормон; П — прогестерон; Э — эстрогены; А — андрогены; Р — релаксин; И — ингибин; ЛГ — лютеинизирующий гормон.
В передней доле гипофиза синтезируются также тиреотропный (ТТГ) и адренокортикотропный (АКТГ) гормоны, а также гормон роста.
ФСГ стимулирует рост и созревание фолликулов яичника, способствует образованию рецепторов ФСГ и ЛГ на поверхности гранулезных клеток яичника, увеличивает содержание ароматаз в зреющем фолликуле и, стимулируя процессы ароматизации, способствует превращению андрогенов в эстрогены, стимулирует продукцию ингибина, активина и инсулиноподобного фактора роста-1, которые играют ингибирующую и стимулирующую роль в росте фолликулов.
Л Г стимулирует:
образование андрогенов в тека-клетках;
овуляцию совместно с ФСГ;
ремоделирование гранулезных клеток в процессе лютеинизации;
синтез прогестерона в желтом теле.
Пролактин стимулирует рост молочных желез и лактацию, контролирует секрецию прогестерона желтым телом путем активации образования в них рецепторов к Л Г.
Синтез пролактина аденогипофизом находится под тоническим блокирующим контролем дофамина, или пролактинингибирующего фактора. Ингибиция синтеза пролактина прекращается во время беременности, лактации. Основным стимулятором синтеза пролактина является тиролиберин, синтезируемый в гипоталамусе.
Циклические изменения в гипоталамо-гипофизарной системе и в яичниках взаимосвязаны и моделируются по типу обратной связи.
Выделяют следующие типы обратной связи:
1) "длинная петля" обратной связи — между гормонами яичника и ядрами гипоталамуса; между гормонами яичника и гипофизом;
2)"короткая петля" — между передней долей гипофиза и гипоталамусом;
3)"ультракороткая петля" — между ГТРГ и нервными клетками гипоталамуса.
Взаимосвязь всех указанных структур определяется наличием в них рецепторов к половым гормонам.
У женщины репродуктивного возраста имеется как отрицательная, так и положительная обратная связь между яичниками и гипоталамо-гипофизарной системой. Примером отрицательной обратной связи является усиление выделения ЛГ передней долей гипофиза в ответ на низкий уровень эстрадиола в раннюю фолликулярную фазу цикла. Примером положительной обратной связи является выброс ЛГ в ответ на овуляторный максимум содержания эстрадиола в крови.
О состоянии репродуктивной системы можно судить по оценке тестов функциональной диагностики: базальная температура, симптом зрачка и кариопикнотический индекс.
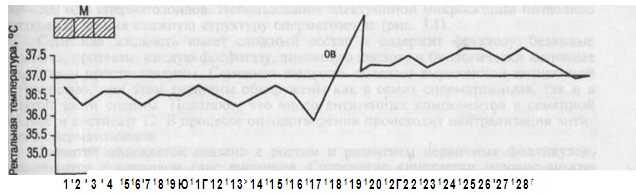
Базальная температура измеряется в прямой кишке утром, до подъема с постели. При овуляторном менструальном цикле базальная температура повышается в лютеиновую фазу цикла на 0,4—0,6 °С и держится в течение всей второй фазы (см. рис.). В день менструации или за день до нее базальная температура снижается. При беременности повышение базальной температуры объясняется возбуждением терморегулирующего центра гипоталамуса под влиянием прогестерона.
Рисунок: Ректальная температура при двухфазном цикле. М — менструация; ОВ — овуляция.
Симптом зрачка отражает изменения в слизи шейки матки. Под влиянием эстрогенов в шейке матки накапливается прозрачная стекловидная слизь, что обусловливает расширение наружного отверстия шейки матки. Максимальное количество слизи наблюдается в предовуляторные дни цикла, наружное отверстие становится темным, напоминает зрачок. Во вторую фазу цикла под влиянием прогестерона количество слизи уменьшается или же она полностью исчезает. Слизь имеет глыбчатое строение. Различают 3 степени симптома зрачка: +, ++, +++.
Кариопикнотический индекс. Под влиянием гормонов яичников также происходят циклические изменения в слизистой оболочке влагалища, особенно в его верхней трети. В мазке из влагалища могут встречаться следующие виды клеток плоского многослойного эпителия: а) ороговевающие, б) промежуточные, в) базальные, или атрофические. Клетки первого типа начинают преобладать по мере нарастания секреции яичниками эстрогенов. На основании определения количественных соотношений клеточных элементов можно судить о степени насыщенности организма эстрогенными гормонами или об их недостаточности. Максимальное число ороговевающих клеток выявляется в предовуляторные дни — 80—88 %, в раннюю фазу пролиферации — 20—40 %, в позднюю фазу секреции — 20-25 %.